Fosfor
|
Rostliny přijímají fosfor ve formě aniontů H2PO4-
nebo HPO42-. Příjem P aniontu probíhá za
podmínek, kdy jeho koncentrace v cytoplazmě výrazně převyšuje
obsah P v živném prostředí. Z tohoto důvodu je přijímán
proti koncentračnímu gradientu (aktivně). Příjem P a jeho
akumulace kořeny je spojená s jeho inkorporací na nízkomolekulární
sloučeniny (estery kyseliny fosforečné, fosforylované cukry,
volné nukleotidy). Studie s 32P prokázaly, že primárními
metabolity jsou adenylnukleotidy (hlavně ATP). Příjem a utilizace
fosforu mají kladný vztah k celé řadě metabolických procesů
uskutečňovaných jak v kořenových buňkách, tak v nadzemních
částech rostlin. Intenzita příjmu P je závislá na obsahu kyslíku
v živném prostředí, světle, teplotě (opt. při 20o
C), poměru H2PO4- ku HPO42-
a na přítomnosti Ca2+, NO3- a BO33-.
Příjem P výrazně inhibují také OH- ionty.
Zabudování
anorganického fosfátu (Pi) do organických forem je velmi rychlé,
a 32P se objevil v kořeni klíčních rostlin hrachu již
za 2-3 sekundy v ATP a o něco později v nukleotidech. Po 10 min.
se již více než 80% přijatého fosforu objevuje v organických
vazbách. Stejný atom P může přecházet z jedné sloučeniny do
druhé několikrát, a proto P řadíme mezi vysoce pohyblivé prvky
se značnou reutilizační schopností. Poločas ATP je 2-20 sec., P
esterů 30 sec., P-lipidů 1-5 hod.. V celé rostlině se fosfor vymění
podle některých autorů za 3 dny. Pro transport fosforu z kořenů
do nadzemních částí rostlin se organické formy defosforylují
hydrolytickými fosfatázami, které uvolňují Pi z organických
vazeb. Fosfatázy jsou zvláště ve vakuolách, kde se P hromadí v
anorganické formě. Z vakuol putuje symplastem do xylémů, kde tvoří
až 75% z veškerého P. Zbývající část se transportuje pravděpodobně
ve formě cholinfosfátu. Rychlost P v xylému je asi 200 cm.hod-1.
S
ohledem na charakter organických vazeb fosfátu v buňce je zřejmé,
že jeho příjem je vázán na respiraci. V rostlinách se fosfor
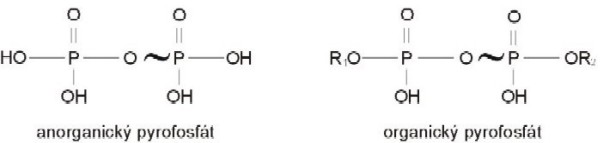
vyskytuje ve formě anorganického nebo organického pyrofosfátu:
Polyfosfáty jsou polymerizované soli typu pyrofosfátu,
které vznikají z ortofosfátu odštěpením vody a představují
rezervní fosfát nacházející se ve vakuolách. Jejich výhodou
je, že se mohou štěpit anorganickými pyrofosfatázami a přitom
uvolňují energii.
Nejpočetnějšími
sloučeninami fosforu jsou jeho organické formy, které v rostlině
tvoří 4 skupiny:
a)
DNA, RNA, P-lipidické, P-estery sacharidů
b)
ATP, ADP, AMP
c)
G-6-P
d)
F-6-P
Fosforylované
sacharidy jsou vlastně labilní meziprodukty metabolismu. Rovněž
labilní jsou i fosfátové vazby v nukleotidech, v kterých se
pyrofosfát váže na sacharidovou složku. Fosfor je složkou dalších
dusíkatých látek (tiaminfosfát, cholinfosfát, fosfoetanolamin),
aktivních látek např. AMP, aktivovaných aminokyselin, sulfátů
aj.. Rezervní organickou formou fosfátů je vícesytný alkohol
fytin. Nachází se v semenech jako Mg- nebo Ca- sůl kyseliny
fytinové.
Experimenty
bylo prokázáno, že při nedostatku fosforu rostlina přestane růst
proto, že vyčerpá Pi z vakuol. Buňka uvolní Pi jen tehdy, má-li
zabezpečený vlastní metabolismus. Proto je nutné, aby byly vytvořeny
pro rostliny vhodné podmínky pro doplňování P v živném prostředí,
a to od klíčení až do fáze jeho maximálního odběru. Funkci
fosfátu v rostlině můžeme rozdělit na energetickou a stavební.
Energetická
funkce je charakterizována schopností ortofosfátu tvořit
esterické vazby (s cukry) o energii s několika kJ/mol P. Proto se
staly anhydrické vazby o-fosfátu snadno mobilní energetickou
konzervou zužitkovatelnou v řadě metabolických reakcí. Na
energetických reakcích se vedle tri- a di-fosfátu adeninu podílí
i inosonové, uridinové, thyamidinové, guanidinové a jiné deriváty.
Podobně se uplatňuje mesoinosit a zvláště hexafosfát, který
neutralizovaný vápníkem a hořčíkem poskytuje velmi stabilní
fytin. Tak i zde cyklická forma cukru váže energii blízkou
hodnotám ATP a ADP, tj. 8-9 kcal (33,4-37,6 kJ).
Pro
rostliny je energeticky nejvýznamnější soustava ADP + P anorg.
Tepelnou stránku těchto procesů je možné považovat za „chladící
směs“, která odebírá energii v centrech intenzivního
energetického metabolismu, a transportuje ji v molekule ATP na jiná
místa, kde ji postupně degraduje po malých kvantech, uvolňuje
teplo a zabraňuje tak tepelnému zničení metabolických center. Tím,
že se může rychle vracet do své minerální ortofosfátové
formy, je v řadě metabolických systémů snadno cyklicky zužitkováván
bez potřeby dalšího fosforu.
Stavební
funkce fosforu vychází prakticky rovněž z tvorby esteru s cukry
a cukernými deriváty. Mezi tyto důležité látky patří mono- a
dinukleotidy, složky enzymů oxidativního charakteru (FMV, FAD,
NAD, NADP, CoA aj.). Další rozsáhlou skupinou jsou fosfoglyceridy,
fosfolipidy, fosfatidy.
Transport
fosfátu v rostlině je spjat s metabolismem a je přímo podmíněn
možností jeho cyklické reutilizace. Jak bylo prokázáno,
pohybuje se fosfát v xylémovém vzestupném proudu převážně ve
formě cukerných esterů. Floémový transport fosfátu je zase výlučně
vázán na jeho organickou formu (ATP, cukerné estery). Minerální
P prochází tedy celým metabolickým procesem než se objeví ve
vodivých pletivech.
Rostliny
potřebují značná kvanta P již v počátečních stadiích růstu.
Potřebný fosfor získávají z fytinu v semeni a dále z lehce přístupných
forem P sloučenin z vnějšího prostředí. V této fázi růstu
kořenový systém není ještě plně rozvinut, a proto má velký
význam hladina přijatelného P v blízkosti primárních kořenů.
Výsledky řady pokusů prokazují významný vliv dodaného fosforu
na zvýšení jeho obsahu v rostlině, což se projevuje v optimálním
růstu rostlin a tvorbě semen se zvýšeným jeho obsahem. Dobrá zásoba
fosforu v půdě zabezpečuje nejen kvalitní výnosy, ale zajišťuje
i jeho koloběh v celém potravním řetězci.
Fosfát
je přítomný v kořenech, stéblech a listech v anorganické formě.
Rozdíly mezi anorg. P a celkovým P jsou největší v mladých
listech, které obsahují relativně větší kvanta org. P ve formě
nukleových kyselin. Vliv
P výživy na obsah P frakcí v listech tabáku (Kakic 1969)
Význam fosforu pro pšenici (doplňkový text) |
||||||||||||||||||||||||||||||||||||||
|
autor textu: Prof. Ing. Rostislav Richter, DrSc. |
||||||||||||||||||||||||||||||||||||||
Poslední aktualizace: 27.01.2004 15:11
Ústav agrochemie a výživy rostlin, MZLU v Brně, Zemědělská 1, 613 00 Brno