Živinný režim půd
|
| makroelementy v půdě | mikroelementy v půdě | užitečné prvky | těžké kovy v půdě | |
||
|
| dusík | fosfor | draslík | vápník | hořčík | síra |
Prvotním
zdrojem půdního dusíku je atmosféra obsahující 77,5 dílů N
(78,08% objemových) převážně ve formě elementárního plynného
dusíku (N2). Vedle toho je součástí atmosféry i řada
oxidu dusíku (NOX) a v malé míře také dusík čpavkový.
Plynný N2 i když se nachází v atmosféře i v půdním
vzduchu, není bez předchozí ionizace přijatelný. Jednou z možností
je elektrický výboj při bouřce, kdy se oxiduje N2 na
NOX, případně až na kyselinu dusičnou. N2
+
O2
----------> 2 NO
2 NO
+
O2
-------->
2 NO2 4 NO2
+
2 H2O
+
O2
---------->
4 HNO3 Na
základě uvedených reakcí přechází do půdy každoročně asi
10-40 kg.ha-1 N.
Dalším
významným zdrojem dusíku je fixace vzdušného N2.
Vlastní redukce vzdušného dusíku vyžaduje značné množství
energie (28 ATP), které představuje přibližně polovinu energie
spotřebované při výrobě N hnojiv (29,3 MJ). Vytvořený amoniak
je vázán na oxokyseliny za vzniku aminokyselin (glutamová,
glutamin). Fixaci vzdušného dusíku rozlišujeme volnou
a symbiotickou. Volnou
fixací se každý rok obohatí ha o 3-12 kg N (v průměru podle půdních
podmínek 5-6 kg). Symbiotickou fixací se u bobovitých váže na
ha 50 - 120 kg N u luskovin, u vojtěšky a jetele 200-300 kg, výjimečně
i více.
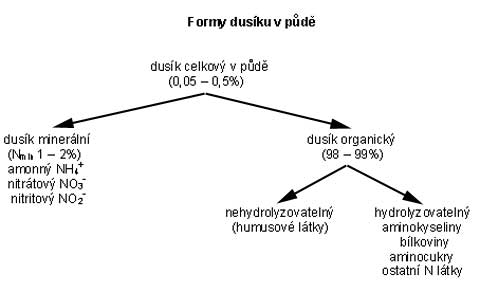
Celkový
obsah dusíku v půdách je velmi rozdílný a kolísá nejčastěji
od 0,05-0,5%.
V orniční vrstvě převážné části půd ČR je 0,1-0,2%
veškerého dusíku. 98 až 99% veškerého N v ornici je přítomno
ve formě organické, zbytek ve formě minerální. Formy N v půdě (IVANIČ et al. 1984)
Obsah celkového dusíku v půdě je hodnotou poměrně stálou,
poněvadž je tvořen sloučeninami těžce chemicky i
mikrobiologicky rozložitelnými. N je zde vázán na aromatická jádra
huminových kyselin, fulvokyselin a huminů. Z tohoto důvodu se
obsah celkového N v půdě často dává do vztahu Cox
a vyjadřuje se poměrem C:N. V našich půdách je uváděná průměrná
hodnota C:N 10-12:1, i když za dostatečné zásobení rostlin dusíkem
považují mnozí autoři ještě poměr 15-18:1. Poměr C:N s
hloubkou klesá, a proto v podorniční vrstvě ho bývá 5-10x méně.
Organické
dusíkaté látky hydrolyzovatelné jsou v půdě mineralizovány až
na amoniak. Rovněž rostlinná a živočišná bílkovinná látka
je pod vlivem proteolytických enzymů vylučovaných různými
skupinami mikroorganismů aerobního a anaerobního charakteru přeměňovaná
postupně přes polypeptidy na peptidy, aminokyseliny a působením
deamináz až na NH3. Do této činnosti jsou zapojeny
jak bakterie (Bact. vulgare, subtilis, mezentericus aj.), tak plísně
(Penicillium, Aspergillus aj.). Uvedenými reakcemi je zajišťováno
spojení mezi organickými N sloučeninami a N minerálním v půdě.
Přitom tyto procesy zajišťují jak rozklad N organických látek,
tak vlivem půdního fyto a zooedafonu i syntézu nových dusíkatých
organických látek. Pro tyto procesy je nutné zajistit pravidelný
přísun organické hmoty. Rychlost mineralizace organického dusíku
na dusík minerální je stimulována celou řadou povětrnostních
a půdních podmínek (teplota, vlhkost, pH, aerace, obsah org. látek
aj.).
Amoniakální
dusík se v půdě nachází v různém stavu (NH4+,
NH3). V nepatrném množství je rozpuštěn v půdním
roztoku ve formě amonných solí, odkud jej mohou rostliny bezprostředně
využít. Část NH4+ je ve výměnné formě
a po jeho vytěsnění z [VSK]
může být rostlinami rovněž přijímán. Část NH3
u lehkých a alkalických zemin může volatilizovat (těkat). Ztráty
na tomto dusíku činí v průměru 20-30% z dodaného dusíku.
Rozpustný
a výměnný NH4+ se může stát nevýměnným
„fixací“ do krystalové mřížky některých minerálů. Illit
poutá více NH4+ než mnotmorillonit a ten
opět více než kaolinit. Množství takto „fixovaného“ NH4+
může být značné. Domácí výsledky z posledních let však
ukazují, že význam tohoto jevu pro dusíkatou výživu rostlin je
minimální, i když má značný ekologický dopad. Část amoniakálního
dusíku je v půdě imobilizována biologickou sorpcí, jejíž
rychlost závisí na intenzitě mikrobiální činnosti půdy. V
biologicky činných půdách podléhá NH4+
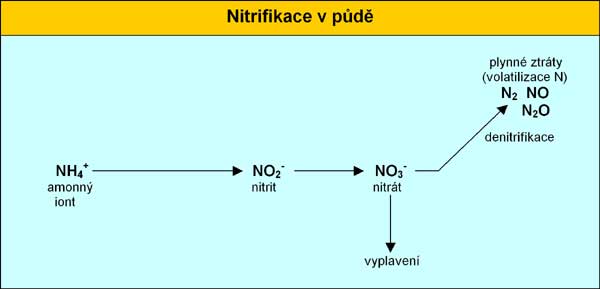
dusík nitrifikaci.
Nitrifikační bakterie získávají z amonných solí potřebnou
energii nezbytnou pro syntézu organických látek a současně jsou
tyto sloučeniny pro ně také zdrojem dusíku. Nitrifikace probíhá
ve dvou stupních. 1.
stupeň - nitritace:
2 NH4+
+
3 O2
---------->
2 HNO2
+
2 H2O
+
2 H+
+
661 J 2.
stupeň - nitratace:
2 HNO2
+
O2
------------>
2 HNO3
+
201 J
Na
oxidaci amonných solí v půdě v 1. stupni se zúčastňují
aerobní baktérie (Nitrosomonas, Nitrosocystis, Nitrosospira aj.),
na oxidaci nitritů (2. stupeň nitrifikace) se podílejí baktérie
rodu Nitrobacter. Při nitrifikaci uvolněné H+
okyselují půdu. Vznikající kyselina dusičná je neutralizována
bázemi sorpčního komplexu, nebo půdního roztoku. Průběh
tohoto biochemického procesu je oblivňován řadou podmínek. Při
dostatečném provzdušení půdy, teplotě 15-30oC,
vlhkosti 40-60% MVK a pH 6,2-9,2, probíhá nitrifikace velmi intenzívně.
Při teplotě 5-10oC je velmi nízká a při teplotách <
5oC se prakticky zastavuje. Během roku se intenzita
nitrifikace mění v závislosti se změnami podmínek. Nejvyšší
intenzity dosahuje v období duben - květen. Příjem dusíku
rostlinami postupnou redukcí nitrifikace obsah dusičnanů i
amoniakálního dusíku v půdě snižuje na relativně stabilní
hodnotu (6.-8. měsíc), aby znovu na podzim dosáhl druhého
maxima. Nitrifikace je dávána do souvislosti s půdní úrodností.
Vysoká intenzita nitrifikace může vést ke značným ztátám dusíku
vyplavením případně posléze i k denitrifikaci. Vzhledem k těmto
skutečnostem se snažíme omezit a usměrnit nitrifikaci použitím
inhibitoru nitrifikace, a tím snížit ztráty nitrátů vyplavením
a omezit jejich kumulaci v rostlinných pletivech.
Ke
ztrátám nitrátového dusíku za současné spotřeby organických
látek dochází hlavně činností denitrifikačních bakterií (Bact.
denitrificans) na oxidy dusíku (NOx) nebo až na elementární
N2. Mikrobiální denitrifikace probíhá intenzívně v
neutrálním až alkalickém prostředí při nedostatku vzduchu.
Podmínkou je dostatek organické hmoty. Průběh reakce lze vyjádřit
sumární rovnicí:
C6H12O6
+
4NO3
----------->
6CO2
+
6H2O
+
2N2
Na redukci NO3- --->
NO2- se podílí enzym nitrátreduktáza, na
redukci NO2- --->
N2 nitritreduktáza. Redukci podporuje přítomnost Mn a
Zn. Vedle denitrifikace biolgické probíhá i denitrifikace chemická.
Při ní kyselina dusitá reaguje s amninokyselinami, aminy, amidy
nebo močovinou až na elementární dusík. Chemická denitrifikace
je intezívnější v kyselém prostředí (pH <
5,5).
CO(NH2)2
+
2 HNO2
----------->
CO2
+
3 H2O
+
2 N2
K
vyplavení dusíku z půdy dochází v závislosti na druhu půdy,
úrovni srážek a na způsobu využití půdy v rozmezí 1,0-54,0
kg.ha-1 N. Pokud se nitrátový dusík dostane mimo kořenovou
zónu (0,8-1 m) je denitrifikace jediný způsob snížení obsahu
nitrátů v podpovrchových vodách. Denitrifikací se ztrácí ročně
v průměru až 8% mineralizovatelného půdního dusíku, a až 20%
N z hnojiv o závislosti na celkové dávce dodaného N.
Vzhledem
k tomu, že chemická denitrifikace může vést k vysokým ztrátám
na drahém dusíku, je třeba agrotechnickými a hnojařskými opatřeními
omezit její intenzitu na nejmenší míru. Dosáhneme toho tím, že
převážnou část dusíku budeme aplikovat ve vegetačním období
a zvýšíme možnost biologické fixace dusíku pěstováním
meziplodin na zelené hnojení nebo zaoráním slámy.
Z
přehledu o půdním dusíku můžeme shrnout, že organické látky
v půdě podléhají mineralizaci, jejímž výsledným prokuktem je
amoniak, který může být sorpčně poután, z půdního roztoku,
přijímán rostlinami, nitrifikován nebo denitrifikován. Za předpokladu,
že je v půdě dostatek organických látek s širokým poměrem
C:N, může být dusík minerální imobilizován. Schéma „turnover
N“ je uvedeno na obrázku. Nulový turnover dusíku
charakterizuje vnitřní cyklus N v půdě, kdy půda je v dynamické
rovnováze, tj. navenek vykazuje nulovou mineralizaci i imobilizaci.
Jestliže převládá mineralizace nad imobilizací obsah Nmin
se v prostředí zvyšuje. Naopak, pokud mineralizace je menší než
imobilizace, N se spotřebovává a může dojít případně až k
dusíkové depresi.
Z dynamiky přeměn půdního dusíku je zřejmé, že přeměna
dusíkatých látek v půdě do forem pro rostliny přijatelných je
závislá na půdních a povětrnostních podmínkách, a proto jsou
značné problémy s využitím Nmin a N lehce
mineralizovatelného N pro praktickou výživu rostlin. Přesto je z
ekonomického a ekologického hlediska výhodné upřesňovat výživu
rostlin touto živinou alespoň podle Nmin a to nejen z
orniční vrstvy, ale i do hloubky 0-0,6 m. Podle obsahu N v půdě
lze pak upravit základní dávku N případně korigovat i dávku N
nutnou k přihnojení během vegetace. |
||
|
autor textu: Prof. Ing. Rostislav Richter, DrSc. |
||
Poslední
aktualizace: 16.01.2007 17:15
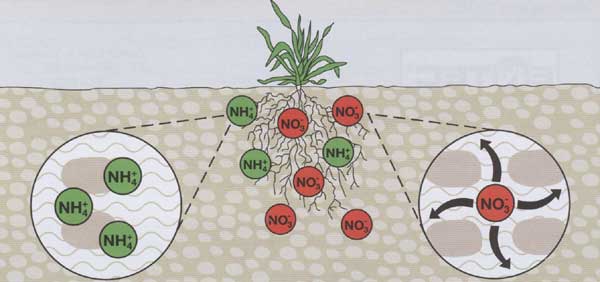
Amonný a nitrátový dusík v půdě |
 Formy dusíku v půdě |
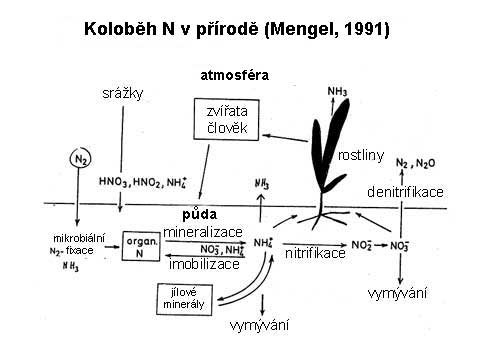
Koloběh dusíku v přírodě |
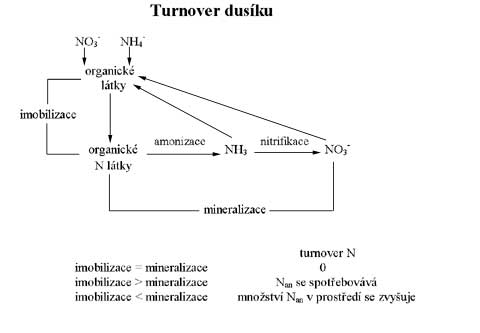
Turnover dusíku |
Ústav agrochemie a výživy rostlin, MZLU v Brně, Zemědělská 1, 613 00 Brno