Příjem živin kořeny
| mechanismus příjmu a transportu látek v rostlině | transport na krátké vzdálenosti | transport na střední vzdálenosti | dálkový transport | |
||
|
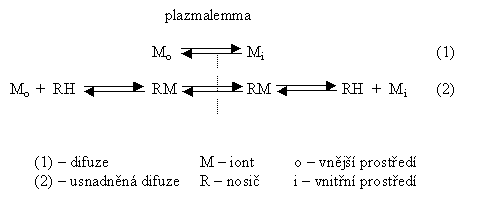
Látky z vnějšího prostředí, aby byly rostlinou metabolicky využity, musí překročit rozhraní mezi vnějším a vnitřním prostředím. Toto rozhraní není vždy totožné s morfologickým povrchem rostliny - jejich orgánů: základní bariérou (o) a vnitřním (i) systémem je plazmalemma. Přes ni vede primární vstup do buněčného prostředí. Příjem látek je spojen s transportem, který překonává prostorové vzdálenosti od místa zdroje látky k místu spotřeby. Podle toho rozlišujeme transport na krátké vzdálenosti (vstup do buňky), na střední a na dlouhé vzdálenosti. Transport je vlastním procesem přemísťování látek a energie. Přechod látky přes jednotkovou plochu rozhraní, daný množstvím převedené látky za čas označujeme jako tok a platí pro něj vztah: J = L . X, kde: L ... vodivost prostředí pro transportovanou látku, X ... příslušná hnací síla. Analyzujeme-li hnací sílu transportu (X), pak můžeme rozlišit děje: a) kde se na její tvorbě podílí např. kinetická energie tepelného pohybu molekul, výpar, gravitační síly, síly vzlínavosti v kapilárních prostorách a ty formy pohybu prostředí, pro něž není hnací silou metabolický proces. V těchto případech mluvíme o dějích pasivních. b) naproti tomu případy, kde X je produktem metabolických procesů, jsou děje považovány za aktivní. Tyto procesy mají svůj původ v samotném buněčném metabolismu, který vytváří energetické i látkové nerovnováhy (gradienty koncentrací), a ty jsou pak vlastní hnací silou aktivních transportních procesů. Snížení metabolické aktivity (např. aplikací inhibitorů, snížení teploty aj.) vede k poruše transportních procesů - ať přímo na membráně nebo nepřímo porušením aktivního konvekčního toku.
Pasivním transportem
je především difúze,
která je nejobecnější formou transportu. Zdrojem energie je
tepelný pohyb molekul (při stejné energii odpovídá rychlost
pohybu molekul (v) různé hmotnosti (m) vztahu: v = k .
Formy pasivního transportu
Procházejí-li ionty strukturami svázanými náboji (většinou zápornými), pak je transport kationtů urychlován, aniontů brzděn. Ionty opačných znamének však nejsou nikdy transportovány nezávisle, ale v roztoku se elektricky vyvažují. Pohyb hydrofilních a iontově aktivních látek přes plazmalemmu umožňuje také usnadněná difúze: látka snadno pronikající lipidní dvojvrstvou membrány se na jedné straně (vnější) spojí s přenášeným iontem a vytvoří komplexní strukturu chelátu. Z něj se uvnitř zase uvolní výměnou za protony nebo hydroxylové ionty. Oba difúzní procesy samy o sobě jsou nezávislé na metabolické energii nebo na jiných zdrojích energie buňky. K typu pasivního transportu možno řadit i transport látek konvekčním tokem prostředí, nezávisle na tom, jakými silami je konvekční tok způsoben. V tomto smyslu platí, že pohybem prostředí není ovlivněn vlastní difúzní pohyb látky.
Aktivní transport
původně označoval transmembránový transport, využívá speciálních
iontově selektivních přenašečů za spotřeby makroergické
vazby (ATP ev. jiné formy) (obr. 2.3 a 2.4). Takový děj nabývá
nutně povahy spřažené enzymatické reakce, jejíž kinetika
vyhovuje enzymatické reakci prvého řádu (je-li ATP v nadbytku): S ohledem na současný (protisměrný) tok H+ a jejich vylučování mohou být tyto přenašeče aktivovány činností protonové pumpy. Podle současného stavu znalostí možno rozlišit na rostlinných membránách několik základních typů transportních systémů:
Pro uniport H+ je popisován velmi účinný systém, tzv. protonová pumpa, která, jak již bylo uvedeno, může aktivovat řadu dalších toků. Transport může být popisován jak z vnějšku dovnitř tak zevnitř ven. Transport protonů H+ běží z vnitřního prostředí do vnějšího a proti němu jde transport ostatních kationtů (K+, Ca2+ aj.), dokud nedojde k rovnováze. Přenos způsoben přenašečem
Rovnováhy jsou dvojího typu: elektrické a koncentrační. Obě se vyrovnávají pasivním pohybem. Jsou však i nerovnováhy mezi vnějším a vnitřním prostředím, které se vyrovnávají aktivním procesem. |
||
|
autor textu: Prof. Ing. Rostislav Richter, DrSc. |
||

Poslední aktualizace: 23.01.2004 09:26
Ústav agrochemie a výživy rostlin, MZLU v Brně, Zemědělská 1, 613 00 Brno