vodík, kyslík, uhlík
|
Tyto prvky tvoří skupinu lehkých biogenních prvků
vzájemně dobře slučitelných. Uhlíkaté skelety tvoří hlavní
podíl sušiny živé hmoty (95% i více u vegetativních orgánů).
V různých fázích vývinu rostlin se tato hodnota mění a určuje
vztah mezi základními chemickými složkami rostlinného těla
(N/C, P/C aj.).
Vodík se vyskytuje jako vysoce mobilní proton (H+) nebo
je vázán kovalentně ve vodě a všech organických sloučeninách.
Kyslík vstupuje spolu s uhlíkem do organických látek
v molekule CO2, další cestou je adice OH- při
hydrolýze, fosforolýze aj. V kořenech jeho nedostaek výrazně
omezuje fyziologické pochody, snižuje příjem vody a živin,
omezuje růst kořenů.
Uhlík ve formě CO2 je hlavním zdrojem veškerého
organického světa na Zemi. Vegetace naší planety spotřebuje ročně
asi 8% veškerého CO2 ze vzduchu, což představuje v průměru
150-220 kg CO2 na ha. Cesty utilizace uhlíku jsou cesty
fotosyntetické přeměny uhlíku a metabolických přeměn. Jejich
funkce se mnohokrát mění během života rostliny. Dynamika
uhlíku je velmi významná pro tvorbu živé hmoty.
Cesty utilizace a přeměn uhlíku jsou biologií
fotoautotrofní rostliny. Uhlík je přijímán ve formě CO2
průduchy listů a v malém množství i kořeny jako aniont HCO3-.
V průběhu fotosyntézy (fáze světelná) je světelná energie
absorbována v chloroplastech a přijímána molekulami chlorofylu a
dalších aktivních pigmentů na ADP z něhož vzniká energeticky
bohatší ATP. Ve druhé fázi dochází k vlastní vazbě CO2
do organických sloučenin. Tato reakce probíhá ve tmě a
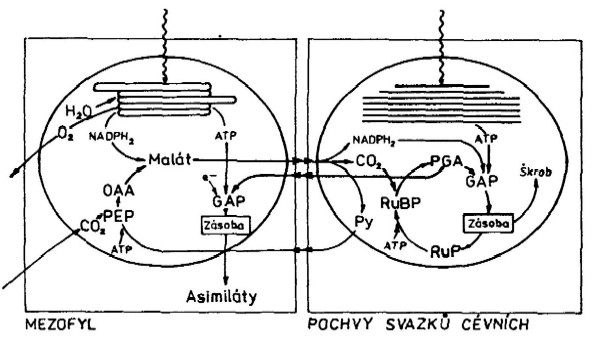
vlastním akceptorem CO2 je u rostlin typu C3 ribulosa-difosfát (pentosa), ze kterého po
reakci vzniknou 2 molekuly 3-fosfoglycerátu (podle Calvinova
cyklu). 3-fosfoglycerát je redukován za účasti ATP na
glyceraldehydfosfát, který je dále po odštěpení kyseliny
fosforečné syntetizován do cukrů (monosacharidy až
polysacharidy).
U řady rostlin (kukuřice, cukrová třtina, proso,
tropické trávy) je akceptorem kyselina oxaloctová, dikarboxylová
kyselina se čtyřmi atomy uhlíku. Tato forma zabudování CO2
byla nazvána C4
cesta. Redukce na uhlíkaté sloučeniny C3 cestou
probíhá u C4 rostlin také, ale příjem CO2
a následné procesy jsou umístěny v prostorově oddělených a
anatomicky odlišných pletivech. V buňkách mezofylu je CO2
vázán na fosfoenolpyruvát (PEP). Tento proces katalyzuje PEP-karboxyláza,
která je velmi účinná při nízkých koncentracích CO2
(i pod 10 µl.l-1).
Pro C3 rostliny je limitující koncentrací tzv.
kompenzační koncentrace, která činí 30-70 µl.l-1.
Z tohoto důvodu zpravidla dosahují C4 rostliny vyšších
rychlostí fotosyntézy než C3 rostliny.
Karboxylace PEP poskytuje kyselinu octovou, která je přeměňována
na kyselinu asparagovou (aspartát) nebo na kyselinu jablečnou (malát).
Žádné z těchto kyselin nemohou být přeměňovány v buňkách
mezofylu, ale musí být přeneseny do buněk pochev svazků
cévních (viz obrázek).
V chloroplastech buněk pochev svazků cévních jsou malát
a aspartát rozštěpeny na CO2 a pyruvát specifickými
enzymy. Uvolněný CO2 je zachycován RuBP a vstupuje do
Calvinova cyklu, zatímco pyruvát se vrací do buněk mezofylu, kde
může sloužit k regeneraci PEP. Tato jednoznačně složitá
kombinace syntézy dikarbonových kyselin a C3-cyklu
dává C4 rostlinám výhodu optimálního využívání
CO2. Velmi vysoká afinita PEP-karboxylázy k CO2
a zvláštní anatomie listů umožňující rostlině ihned opětovně
využít CO2 uvolněný při fotorespiraci. C4
rostliny jsou proto schopny využívat i nejvyšší intenzity světla
a poskytují vysoké výtěžky fotosyntézy, které se projeví
mimo jiné rychlejším rozdělením produktů asimilace a vysokou
produkci sušiny. Zvýšení obsahu CO2 zvláště ve sklenících může přinést výrazný efekt právě u rostlin C3, u nichž je saturace CO2 dosaženo až při vyšších koncentracích a u kterých je zvýšení koncentrace CO2 v ovzduší spojeno i s vyšší rychlostí fotosyntézy. Vyšší obsah CO2 může být skleníkovým efektem spojen i se zvýšením teploty. Pro zvýšení intenzity výroby ve sklenících a foliovnících se u zahradních plodin doporučuje zvýšit koncentraci CO2 na 0,6-0,9%, a to v době nejvyšší fotosyntetické aktivity rostlin. Potom je obvykle obnovena jeho běžná koncentrace větráním. |
||
|
autor textu: Prof. Ing. Rostislav Richter, DrSc. |
||
Poslední aktualizace: 23.01.2004 18:25
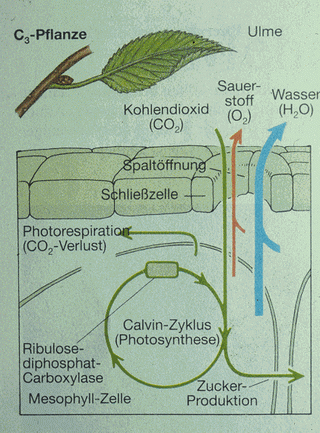
Asimilace uhlíku v C3 rostlině (jilm) |
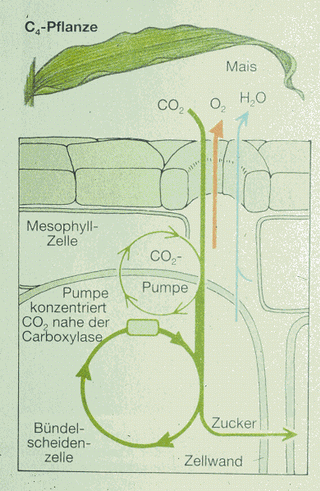 Asimilace uhlíku v C4 rostlině (kukuřice) |
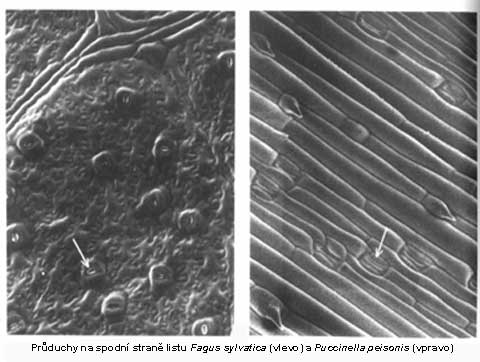
Mikroskopický snímek spodní strany listů |
Ústav agrochemie a výživy rostlin, MZLU v Brně, Zemědělská 1, 613 00 Brno